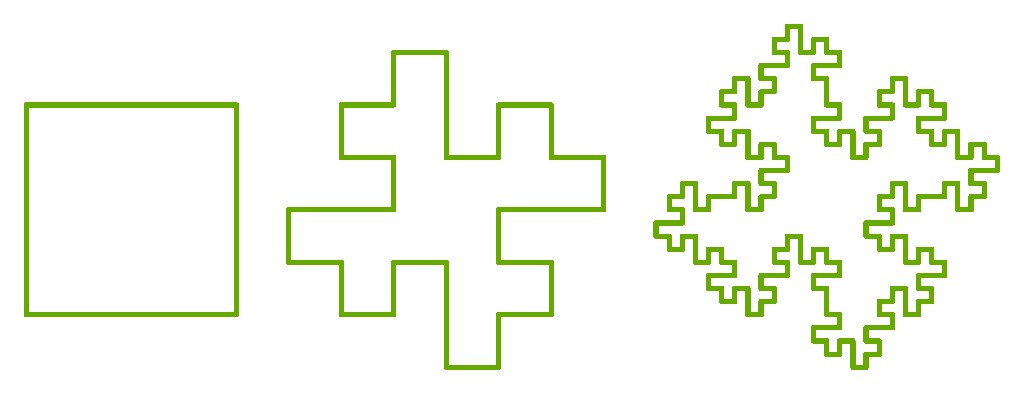
Even Mandelbrot never seemed to ask himself what the distinct design advantage of fractalic structure might be: like most mathematicians, he's so impressed by the elegance of the encoding scheme that every other consideration fades from view. But that simplicity and rigor might actually be a hindrance in organic terms, since an organism may lose too much adaptive range for whatever savings in encoding is gained: it'd be worth asking the question why life didn't develop further along purely fractalic lines, but seemed to abandon it early on in favor of other symmetries.
It strikes me that the primary advantage of the fractal is not its encoding elegance but its amplification potential. Realized fractality acts inevitably as an amplifier of weak and partial signal. Like cracking a whip, the trick is to follow the gentle curve of its unfolding, to induce acceleration without exceeding its accumulating inertia. The cumulative signal grows logarithmically as it proceeds along the fractal scale: amplification response and generative formula coincide.
This assumes that every scaling step, a proportional response grows the signal while matching the antennal sensitivity: such that we don't waste realized surface area for signal transduction. This reminds me of how surfers read waves: they sense which among the series has the best potential to match both the amplifying properties of the shoreline and the ambient perturbations of the medium - if there's too much amplitude early it'll crest before the contours of the seafloor can express themselves.
My guess is that if we plotted the response in a naturally occurring fractal antenna, we'd find it was some proportion of Euler's number, a smooth growth function. The idea is that we don't want turbulence in growth. If we expend too much of the incoming signal - which is nothing but energy - too early in the response, we may fail to explore its full scope. We want a smooth distribution along our growth curve, gaining in amplitude and bandwidth while we eat into our signal budget. From the organic point of view, we don't know the value of any incoming information until it's been fully processed, but some signals are essential, so both efficiency and poised sensitivity are needed: fractal structure seems to allow for the gradual entrainment of some maximum of the network by simply scaling up, from the most responsive smaller scales to the larger less sensitive scales. This should also allow simultaneous signals to compete in parallel. Again during the signal event, neither the duration, nor the amplitude, nor the frequency band, nor the informative value is known until it's traversed.
This assumes that the signal is sufficiently uniform such that fractality has a chance to kick in - to utilize its own self-similar scaling to explore the signal without wasting time nor incoming energy. Imagine what happens when you flick a tuning fork: there's short but intense period of turbulent discharge, in which you've induced useless noise, before enough signal is dissipated that the fork can respond according to its resonant frequency: this is what we want to avoid.
So the question becomes: is this a plausible model for the kind of neural excitation I have in the back of my mind? Namely myoclonus vs the "kundalini shivers":
- The kundalini shivers represent the remnant turbulence of an excursion past the critical point, triggered typically by inhalation.
- Myoclonic cascade represents the fallback, from some supercritical excitation to subcritical, triggered by exhalation.
It's worth emphasizing that these are merely symptoms, not the traversals themselves: these are merely turbulent signatures, vestigial signs. Keeping in mind that most of what happens in the nervous system is invisible because it's efficient: the only way to characterize it is via errata, excess noise, essentially exhaust - which is why overexcitation via psychedelia and deep meditation is so valuable. We need clues to how the smooth transduction event unfolds, the invisible induction and dissipative loss.
There's a caveat that hasn't yet been expressed: criticality may not yield a grand unified theory of neural activity. It may only represent one possible mode: maybe callibration, maybe rare and expensive peak neural performance, it may not even be particularly adaptive to sustain. My own experience is that it's very brief at best. On the other hand, it does seem that we're supposed to experience critical states every waking cycle, that we transition from sleep states of very low excitation to supercritical positions, that we should be more or less always on the move between these extremes, easily rising and gently falling - that would be the profile of a healthy creature. The larger context here is probably the wake-sleep cycle, and I rarely see this considered in all the talk about neural criticality: that we should be considering a continuum from the broadly coherent oscillations of deepest sleep to critical scale-invariant activation.


